
Konrad Zinsmaier, Ph.D.

Rm 627 Gould-Simpson
Research Interests
My laboratory has a long-standing interest in understanding molecular mechanisms that facilitate synaptic function and plasticity, and mechanisms that protect synapses from neurodegeneration. We pursue these goals by undertaking a multidisciplinary approach using genetically modified Drosophila as a model system.
Background
Synapses are specialized cell-cell contact sites to transmit signals between neurons on a sub-millisecond scale. This is achieved by precisely aligning the neurotransmitter release machinery in the presynaptic axon terminal to the neurotransmitter receptor apparatus of the postsynaptic neuron to electrically excite or silence the postsynaptic neuron. The accuracy of this process is vital as even subtle changes in synaptic function can disturb neuronal networks and cause pathological abnormalities that lead to neurological and/or neuropsychiatric disorders.
Synapses face significant operational challenges. Maintaining normal function requires the efficient replacement of aged, overused, and potentially toxic proteins and lipids, which is especially challenging for presynaptic terminals as they are often located remarkably distant from sites of biogenesis in the neuronal cell body. These constraints emphasize the unique need of presynaptic terminals for specialized mechanisms to ensure their long-term operation.
The effect of a synaptically transmitted signal can vary enormously, as it can be modified by various cues including the recent history of activity. Next to adjusting the efficacy of individual synapses, neurons can also change the number of synaptic connections between two neurons either by forming new synapses or eliminating existing ones. Such dynamic rearrangements of synaptic efficacy are termed synaptic plasticity and can last from milliseconds to months.
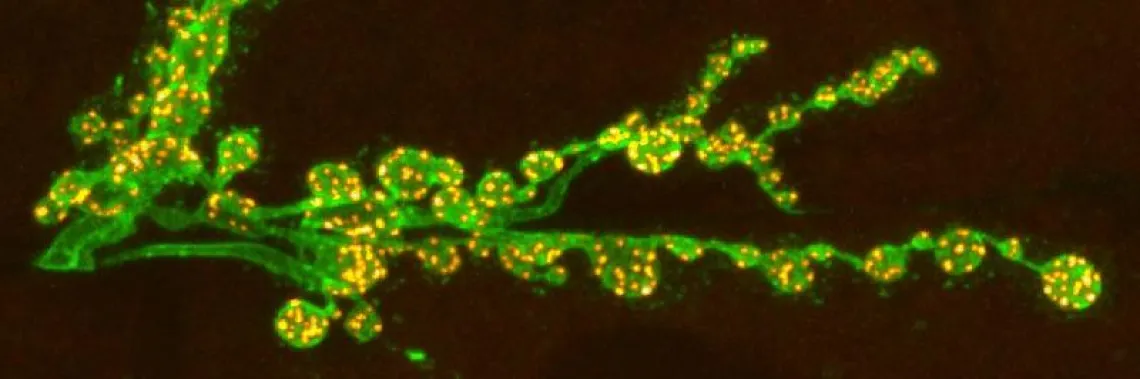
Synaptic plasticity can be divided into three classes: i) short-term plasticity (milliseconds to minutes) allows synapses to perform critical computational functions in neural circuits; ii) long-term plasticity (hours to months or longer) provides the basis for most models of learning and memory, and for developing critical aspects of the cortex; iii) homeostatic plasticity maintains appropriate levels of activity and connectivity within neural networks. All three forms of synaptic plasticity together with others provide the basis for the extraordinary computational and learning abilities of the brain.
Research
The Neuroprotective Role of Cysteine-string Protein
Failure of maintaining operational synapses is likely a major cause for various neurodegenerative diseases. Cysteine-string protein (CSP) has emerged as a critical neuroprotective factor maintaining synaptic function. We and others showed that CSP prevents synaptic failure and neurodegeneration by maintaining the activity of critical synaptic proteins like SNAREs and Dynamin, which mediate SV fusion and recycling. CSP also facilitates the secretion and clearance of misfolded toxic proteins implicated in Huntington’s Disease, Parkinson’s Disease, ALS, and others. Consistently, CSP overexpression in an ALS fly model suppressed synaptic and locomotor defects.
Dominant mutations in human CSP/DNAJC5 cause the neuronal ceroid lipofuscinosis (NCL) ANCL/CLN4, a neurodegenerative lysosomal storage disease. To gain insights into disease mechanisms, we developed two Drosophila models that recapitulate the key pathology of post-mortem CLN4 patient brains. Further analysis revealed a novel link between CLN4 mutant CSP and lysosomal failure. Instead of being localized to SVs, mutant CSP accumulates on abnormal prelysosomes in both axons and somata that likely correlate with abnormal endosomal membrane structures. Genetics analysis showed that both CLN4 alleles act mostly as hypermorphic gain of function mutations increasing one or several functions of CSP. Currently, we are examining molecular mechanisms that cause the prelysosomal failure and neurodegeneration induced by CLN4 mutations.
DCAF12’s role for synaptic function and plasticity.
Synaptic plasticity is conferred by numerous mechanisms altering protein activity, trafficking, localization, turnover, and de novo protein expression. Early studies indicated that changes in pre- and postsynaptic architecture and efficacy can be controlled by ubiquitination, a dynamic and reversible post-translational protein modification, which can regulate protein expression, activity, localization, or degradation.
We identified a novel ubiquitination machinery that mediates synaptic function and plasticity. DCAF12 was biochemically identified as a component of human Cullin 4 (Cul4) E3 ubiquitin ligase complexes, which have been linked to intellectual disability. Our work shows that presynaptic DCAF12 is required for normal nerve-evoked neurotransmitter release and homeostatic synaptic plasticity at fly neuromuscular junctions. In addition, postsynaptic nuclear DCAF12 controls synaptic efficacy by downregulating the synaptic ratio of the glutamate receptor (GluR) subunits GluRIIA/B in a Cul4-dependent manner. Currently, we are exploring targets that underly DCAF12’s role promoting homeostatic synaptic plasticity.
Teaching
NSCS 195B Engaging Topics in Neuroscience and Cognitive Science, Learning & Memory
NROS 430 Neurogenetics
NRSC 695F/G Neuroscience Colloquium and Journal Club
NRSC 588 Principles of Cellular and Molecular Neurobiology
Selected Publications
Zinsmaier, K.E. (2020). Mitochondrial Miro GTPases coordinate mitochondrial and peroxisomal dynamics, Small GTPases, DOI: 10.1080/21541248.2020.1843957.
Imler, E., J.S. Pyon, Y. Zhang, S. Kindelay, M. Torvund, S. Chandra, K.E. Zinsmaier (2019). A Drosophila model of neuronal ceroid lipofuscinosis CLN4 reveals a hypermorphic gain of function mechanism. eLife 8, e46607. DOI: 10.7554/eLife.46607.
Patron, L.A., Nagatomo, K., Eves, D.T., Imad, M., Young, K., Torvund, M., Guo, X., Rogers, G.C., and Zinsmaier, K.E. (2019). Cul4 ubiquitin ligase cofactor DCAF12 promotes neurotransmitter release and homeostatic plasticity. J. Cell Biol. 218, 993–1010. DOI: 10.1083/jcb.201805099.
Parker, S.S, A. Moutal, S. Chandrasekaran, S. Cai, M.R. Roman, A. Koshy, R. Khanna, K.E. Zinsmaier, G. Mouneimne (2018). High fidelity cryopreservation and recovery of primary rodent cortical neurons. eNeuro 5, 1–21. DOI: 10.1523/ENEURO.0135-18.2018.
Coyne, A.N, I. Lorenzini, C.C. Chou, M. Torvund, R. S Rogers, A. Starr, B.L. Zaepfel, J. Levy, J. Johannesmeyer, J.C. Schwartz, H. Nishimune, K.E. Zinsmaier, W. Rossoll, R. Sattler, D.C. Zarnescu (2017). Post-transcriptional inhibition of Hsc70-4/HSPA8 expression leads to synaptic vesicle cycling defects in multiple models of ALS. Cell Reports 21, 110-12. DOI: 10.1016/j.celrep.2017.09.028.
Yong Zhou, C.O. Wong, K. Cho, D. van der Hoeven, H. Liang, D.P. Thakur, J. Luo, M. Babic, K.E. Zinsmaier, M.X. Zhu, H. Hu, K. Venkatachalam and J.F. Hancock (2015). Membrane potential modulates plasma membrane phospholipid dynamics and K-Ras signaling. Science 349, 873. DOI: 10.1126/science.aaa5619.
Babic, M. G.J Russo, A.J. Wellington, R. Sangston, M. Gonzalez, K.E. Zinsmaier (2015). Miro's N-terminal GTPase domain is required for transport of mitochondria into axons and dendrites. J Neurosci. 35, 5754. DOI:10.1523/JNEUROSCI.1035-14.2015.
Imler, E. and K.E. Zinsmaier (2014). TRPV1 Channels: Not So Inactive on the ER. Neuron 84, 659. DOI:10.1016/j.neuron.2014.10.052.
Tsai, P., M.M. Course, J.R. Lovas, C. Hsieh, M. Babic, K.E. Zinsmaier, X.Wang (2014). PINK1-mediated Phosphorylation of Miro Inhibits Synaptic Growth and Protects Dopaminergic Neurons in Drosophila. Scientific Reports 4: 6962. DOI: 10.1038/srep06962.
Liu, Y.C., M.W. Pearce, T. Honda, T.K. Johnson, S. Charlu, K. R. Sharma, M. Imad, R.E. Burke, K.E. Zinsmaier, A. Ray, A. Dahanukar, M. de Bruyne, C.G. Warr (2014). The Drosophila phospholipid flippase dATP8B is required for odorant receptor function. PLOS Genetics 10. DOI: 10.1371/journal.pgen.1004209.
Babic, M. and K.E. Zinsmaier (2011). Memory, synapse stability, and β-Adducin. Neuron, 69:1039. DOI:10.1016/j.neuron.2011.03.004.
Zinsmaier, K.E. and M. Imad (2010). Cysteine-string protein’s role at the synapse. In “Folding for the synapse” (eds. A. Wyttenbach and V. O'Connor), Springer New Yord, pp. 145-176.
Zinsmaier KE (2010). Cysteine-string protein's neuroprotective role. J Neurogenet 24:120-132. DOI:10.3109/01677063.2010.489625.
Chouhan, A.K. J. Zhang, K.E. Zinsmaier and G.T. Macleod (2010). Presynaptic mitochondria in functionally different motor neurons exhibit similar affinities for Ca2+ uptake but exert little influence as Ca2+ buffers at nerve firing rates in situ. J. Neuroscience 30, 1869-81. DOI: 10.1523/JNEUROSCI.4701-09.2010.
Zinsmaier KE, Babic M, Russo GJ (2009). Mitochondrial transport dynamics in axons and dendrites. Results Probl Cell Differ 48, 107-139. DOI: 10.1007/400_2009_20. (Book version: In “Cell Biology of the Axon”, E. Koenig, (ed.). Heidelberg, Springer).
Cziko, A.J., T.C. McCann, I.C. Howlett, S.A. Barbee, R.P. Duncan, R. Luedemann, D. Zarnescu, K.E. Zinsmaier, R.R. Parker, and M. Ramaswami (2009). Genetic Modifiers of dFMR1 encode RNA-granule components in Drosophila, Genetics 182, 1051-60. DOI: 10.1534/genetics.109.103234.
Zarnescu, D.C. and K. E. Zinsmaier (2009). Ferrying wingless across the synaptic cleft. Cell 139, 229-231. DOI: 10.1016/j.cell.2009.09.032.
Russo, G.J., K. Louie, A. Wellington, G.T. Macleod, F. Hu, S. Panchumarthi, and K.E. Zinsmaier, (2009). DrosophilaMiro is required for both anterograde and retrograde axonal mitochondrial transport. J Neuroscience 29, 5443-5455. DOI: 10.1523/JNEUROSCI.5417-08.2009.
Louie, K., G.J. Russo, D.B. Salkoff, A.Wellington, and K.E. Zinsmaier (2008) Effects of imaging conditions on mitochondrial transport and length in larval motor axons of Drosophila. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 151, 159-172. DOI: 10.1016/j.cbpa.2008.06.023.
Dawson-Scully, K, Y. Lin, M. Imad, J. Zhang, L. Marin, J.A. Horne, I.A. Meinertzhagen, S. Karunanithi, K.E. Zinsmaier and H.L. Atwood (2007). Morphological and Functional Effects of Altered Cysteine String Protein at the Drosophila Larval Neuromuscular Junction. Synapse 61, 1-16. DOI: 10.1002/syn.20335.
Macleod, G.T., and K.E. Zinsmaier (2006) Synaptic homeostasis on the fast-track. Neuron 52, 569-571. DOI: 10.1016/j.neuron.2006.11.006.
Guo, X., G.T. Macleod, A. Wellington, F. Hu, J. Zhang, S. Panchumarthi, M. Schoenfield, L. Marin, M.P. Charlton, H. L. Atwood, and K. E. Zinsmaier (2005). The GTPase dMiro is required for axonal transport of mitochondria to Drosophila synapses. Neuron 47, 379-393. DOI: 10.1016/j.neuron.2005.06.027.
Bronk, P., Z. Nie, M.K. Klose, K. Dawson-Scully, J. Zhang, R.M. Roberston, H. L. Atwood, and K. E. Zinsmaier. (2005) The multiple functions of cysteine-string protein analyzed at Drosophila nerve terminals. J Neuroscience 25, 2204-2214. DOI: 10.1523/JNEUROSCI.3610-04.2005.
Song, W. and K. E. Zinsmaier (2003). Endophilin and Synaptojanin hook up to promote synaptic vesicle exocytosis. Neuron 40, 665-667. DOI: 10.1016/S0896-6273(03)00726-8.
Song, W., R. Ranjan, K. Dawson-Scully, P. Bronk, L. Marin, L. Seroude, Y. Lin, Z. Nie, H. L. Atwood, S. Benzer, and K. E. Zinsmaier. (2002) Presynaptic regulation of neurotransmission in Drosophila by the G protein-coupled receptor Methuselah. Neuron 36, 105-119. DOI: 10.1016/S0896-6273(02)00932-7.
Zinsmaier, K.E. & P. Bronk. (2001). Molecular chaperones and the regulation of neurotransmitter exocytosis. Biochemical Pharmacology 62, 1-11. DOI: 10.1016/S0006-2952(01)00648-7.
Bronk, P., J. J. Wenniger, K. Dawson-Scully, X. Guo, S. Hong, H. L. Atwood, and K. E. Zinsmaier (2001). DrosophilaHsc70-4 is critical for neurotransmitter exocytosis in vivo. Neuron 30, 475-488. DOI: 10.1016/S0896-6273(01)00292-6.
Dawson-Scully, K., P. Bronk, H. L. Atwood, and K. E. Zinsmaier (2000). Cysteine-string protein increases the calcium-sensitivity of neurotransmitter exocytosis in Drosophila. J. Neuroscience 20, 6039-6047. DOI: 10.1523/JNEUROSCI.20-16-06039.2000.
Nie, Z., R. Ranjan, J. Wenniger J., S. Hong, P. Bronk, and K. E. Zinsmaier. (1999) Overexpression of cysteine-string protein in Drosophila reveals interactions with syntaxin. J. Neuroscience 19: 10270-10279. DOI: 10.1523/JNEUROSCI.19-23-10270.1999.
Eberle K. K., K. E. Zinsmaier, S. Buchner, M. Gruhn, M. Jenni, C. Arnold, C. Leibold, D. Reisch, N. Walter, E. Hafen, A. Hofbauer, G.O. Pflugfelder, E. Buchner (1998) Wide distribution of the cysteine string proteins in Drosophila tissues revealed by targeted mutagenesis. Cell Tiss. Res. 294: 203-217. DOI: 10.1007/s004410051170.
Ranjan, R., P. Bronk, K.E. Zinsmaier (1998). Cysteine string protein is required for calcium secretion coupling of evoked neurotransmitter release, but not for vesicle vesicle recycling. J. Neuroscience 18: 956-964. DOI: 10.1523/JNEUROSCI.18-03-00956.1998.
Umbach, J.A., K.E. Zinsmaier, K.K. Eberle, E. Buchner, S. Benzer, and C. B. Gundersen (1994) Presynaptic dysfunction in Drosophila csp mutants. Neuron 13: 899-907. DOI: 10.1016/0896-6273(94)90255-0.
Zinsmaier, K.E., K.K. Eberle, E. Buchner, Walter, N., and S. Benzer. (1994) Paralysis and Early Death in Cysteine-String Protein Mutants of Drosophila. Science 263: 977-990. DOI: 10.1126/science.8310297.
Reifegerste, R., S. Grimm, S. Albert, N. Lipski, G. Heimbeck, A. Hofbauer, G.O. Pflugfelder, D. Quack, C. Reichmuth, K.E. Zinsmaier, S. Buchner, and E. Buchner (1993). An invertebrate Calcium-binding Protein of the Calbindin Subfamily: Protein structure, Genomic Organization and Expression Pattern of the Calbindin-32 Gene of Drosophila. J. Neuroscience 13(5): 2186-2198. DOI: 10.1523/JNEUROSCI.13-05-02186.1993.
Zinsmaier, K.E., A. Hofbauer, G. Heimbeck, G.O. Pflugfelder, S. Buchner, E. Buchner (1990). A cysteine-string protein is expressed in retina and brain of Drosophila. J. Neurogenetics 7, 15-29. DOI: 10.3109/01677069009084150.